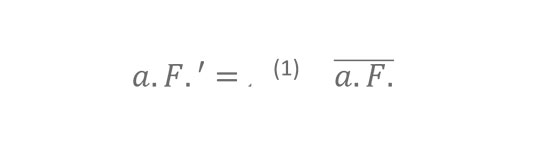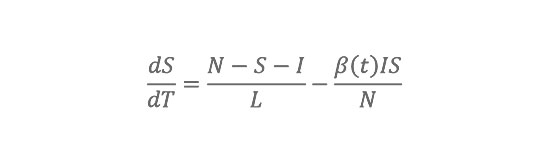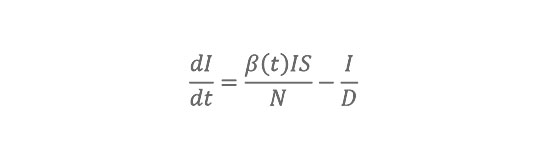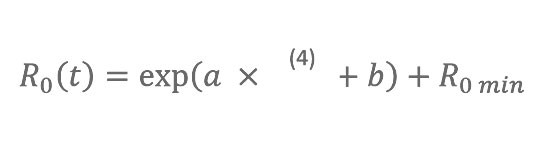Study by Jeffrey Shaman1*, Virginia E. Pitzer2,3,4, Cécile Viboud2, Bryan T. Grenfell2,4,5, Marc Lipsitch6,7,8
In temperate regions, wintertime influenza epidemics are responsible for considerable morbidity and mortality [1]. These seasonal epidemics are maintained by the gradual antigenic drift of surface antigens, which enables the influenza virus to evade host immune response [2]. Recent influenza epidemics have resulted from the cocirculation of three virus (sub)types, A/H1N1, A/H3N2, and B, with one generally predominant locally in a given winter [3–5]. In contrast, influenza pandemic activity can occur any time of year, including during spring or summer months, in the rare instances when a novel virus to which humans have little or no immunity jumps from avian or mammalian hosts into the human population, as in the on-going H1N1v pandemic [6–9]. Despite numerous reports describing wintertime transmission of epidemic influenza in temperate regions [10], our understanding of the mechanisms underlying influenza seasonal variation remains very limited.
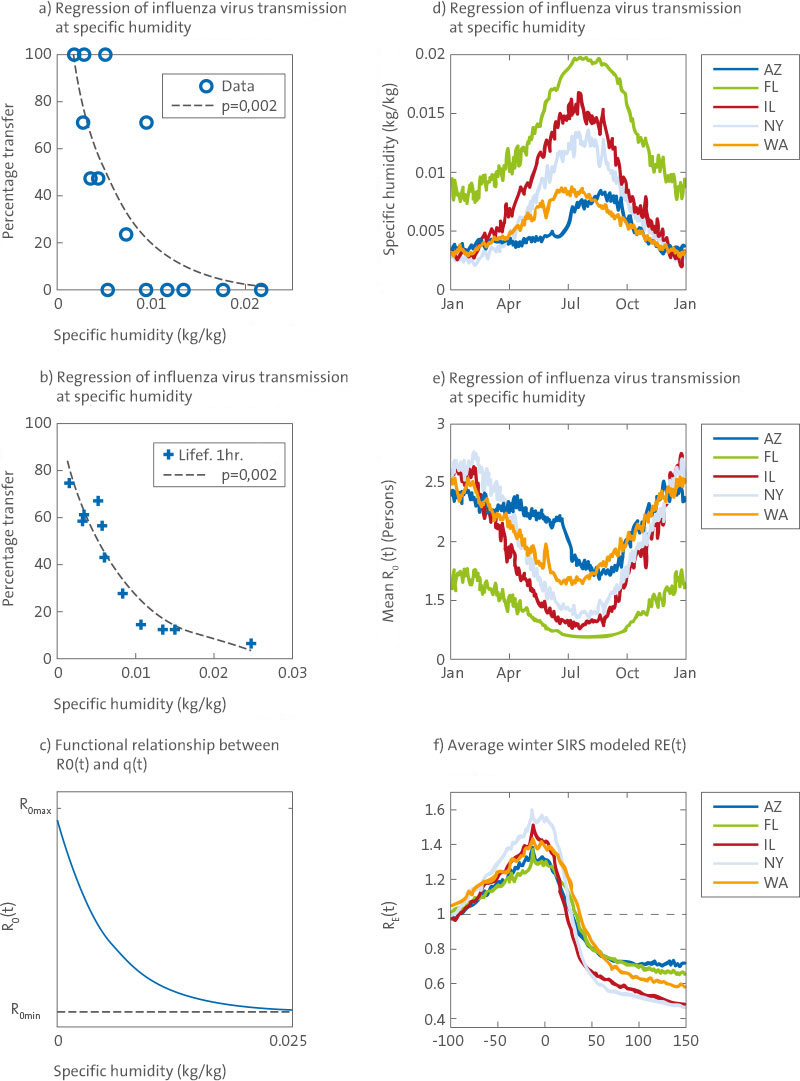
Experimental studies suggest that influenza virus survival within aerosolized droplets is strongly associated with the absolute humidity (AH) of the ambient air, such that virus survival improves markedly as AH levels decrease [11]. A similar relationship is observed between AH and airborne influenza virus transmission among laboratory guinea pigs, in that transmission increases markedly as AH levels decrease (Figure 1).
Figure 1
Analyses of laboratory data, environmental data, and SIRS model simulations. (A) Log-linear regression of guinea pig airborne influenza virus transmission data [14,15] on specific humidity (a measure of AH); (B) log-linear regression of 1-h influenza virus survival data [28] on specific humidity; (C) functional relationship between R
0(t) and q (t) according to equation 4; (D) daily climatology of the reanalysis of the specific humidity 2 m above the ground from 1972-2002 by the NCEP-NCAR [23] for Arizona, Florida, Illinois, New York State and Washington State; (E) daily averages for R
0(t) from 1972–2002, derived from the climatic data on the specific humidity using the best-fit parameter combination from SIRS simulations (R
0max = 3,52; R
0min = 1,12) and the functional form (Figure 1C and Equation 4); (F) average R
E(t) for all winter outbreaks in the ten best-fit simulations in each state, from 100 days before to 150 days after the outbreak began (at least 400 infections / day in the two weeks before; at least 5000 infections / day on at least 1 day in the following 30 days). Figures 1A and 1B are new drawings according to Shaman and Kohn [11] using the specific humidity as a measure of the absolute humidity. doi:10.1371/journal.pbio.1000316.g001
Results
Absolute humidity and the Onset of Wintertime Influenza Outbreaks
In the temperate zones of the world, absolute humidity is lowest in winter and highest in summer (Figure 1D). This seasonal cycle favors an increase in both survival and winter transmission of influenza viruses and may explain the observed seasonal peak of winter influenza morbidity and mortality. Long-term observations of excessive mortality from pneumonia and influenza (P&I) in the United States, which are a robust indicator of the timing and impact of epidemics at the national and local levels, clearly show that winter mortality peaks each year achieved [4] (Figure S1).
Here we are developing an epidemiological underpinning of these previously laboratory-related findings, which imply that the absolute humidity is largely responsible for the seasonal transmission of influenza. First, we analyze the spatial and temporal variation in the occurrence of influenza epidemics for the continental USA between 1972 and 2002 and correlate this observed variability with records of the absolute humidity for the same time periods and locations. Then we show that a mathematical model of influenza transmission in the United States can represent the spatial and temporal variation of epidemic influenza when the daily absolute humidity conditions within each state are used to calculate the base reproduction number, R
0(t), to modulate the influenza virus.
Our first test of the hypothesis that low AH drives wintertime increases of influenza transmission is to assess whether the onset of the influenza epidemic each winter—which shows substantial annual variation (Figure S1)—corresponds to a period of unusually low AH. We define the onset of wintertime influenza as the date at which, for the 2 wk prior, the observed excess P&I mortality rate had been at or above a prescribed threshold level (e.g., 0.01 deaths/100,000 people/day). This onset date was identified separately for each of the 30 winters in the 1972–2002 observational record at each of the 48 contiguous states plus the District of Columbia (DC). We then examined the anomalous AH (AH9) conditions prior to and following these onset dates. AH9 is the local daily deviation of AH from its 31-y mean for each day (as shown for five states in Figure 1D), defined as:
where (
) denotes the 1972–2002 daily average value. At temperate latitudes, such as in the US, wintertime AH levels are already much lower than summer (Figure 1D). By using AH9, we can determine whether the onset of wintertime influenza occurs when AH is above or below typical local daily AH levels.
Negative AH9 values are typically observed beginning 4 wk prior to the onset of influenza epidemics (Figure 2), with the largest excursion occurring 17 d prior to onset. This result is robust to the choice of the mortality threshold level used to define onset date (from 0.001 to 0.02 excess P&I deaths/100,000 people/day). To assess the statistical significance of the association between negative wintertime AH9 and epidemic onset, we bootstrapped the distribution of observed wintertime AH9 records and found strong statistical support (p,0.0005, see Text S1). Depending on the threshold used to define onset, 55%–60% of onset dates demonstrate negative AH9 averaged over the 4 wk prior to onset. Although highly statistically significant, this shift from the expected 50% likelihood is small. These findings indicate that negative AH9 are not necessary for wintertime influenza onset but instead presage an increased likelihood of these onset events. In effect, negative AH9 in the weeks prior to onset provide an additional increase of influenza virus survival and transmission over typical local wintertime levels and may further facilitate the spread of the virus.
Negative AH values are typically observed beginning 4 wk prior to the onset of influenza epidemics (Figure 2), with the largest excursion occurring 17 d prior to onset. This result is robust to the choice of the mortality threshold level used to define onset date (from 0.001 to 0.02 excess P&I deaths/100,000 people/day). To assess the statistical significance of the association between negative wintertime AH9 and epidemic onset, we bootstrapped the distribution of observed wintertime AH9 records and found strong statistical support (p,0.0005, see Text S1). Depending on the threshold used to define onset, 55%–60% of onset dates demonstrate negative AH9 averaged over the 4 wk prior to onset. Although highly statistically significant, this shift from the expected 50% likelihood is small. These findings indicate that negative AH9 are not necessary for wintertime influenza onset but instead presage an increased likelihood of these onset events. In effect, negative AH9 in the weeks prior to onset provide an additional increase of influenza virus survival and transmission over typical local wintertime levels and may further facilitate the spread of the virus.
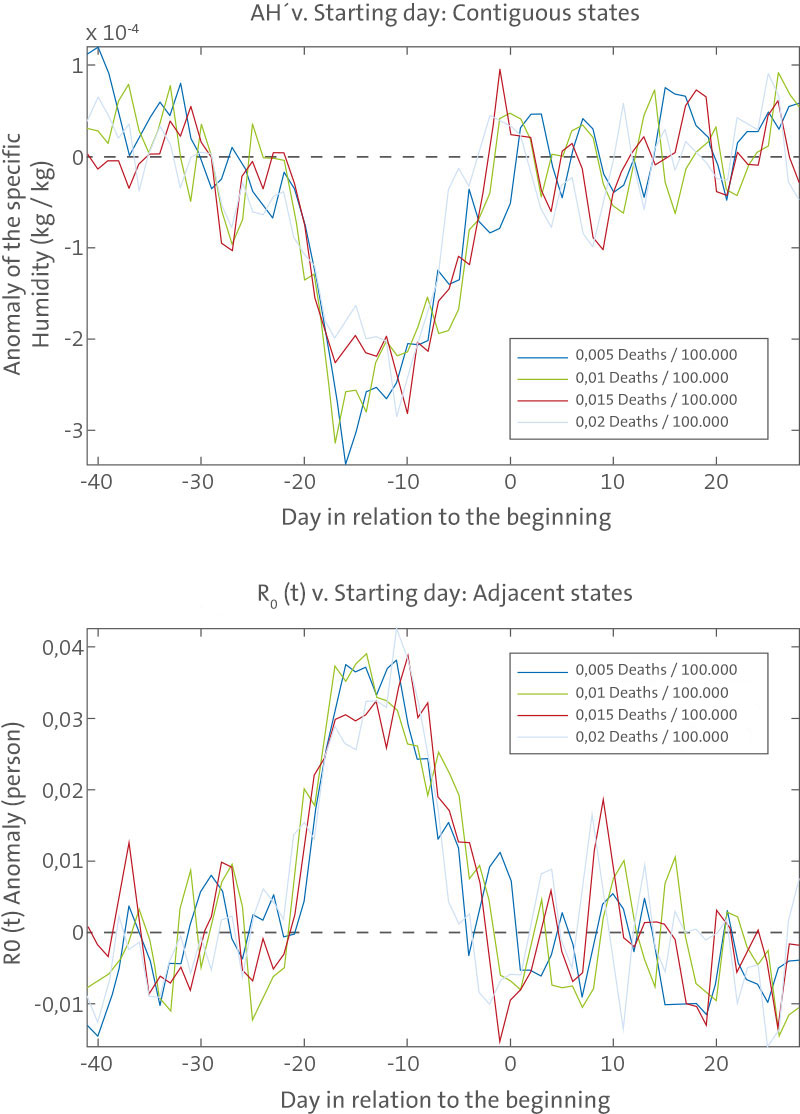
AH9 associated with the observed onset of epidemic influenza. Top, plots of AH9 averaged for the site-winters with an influenza
outbreak showing the 6 wk prior to and 4 wk following outbreak onset. The conditions at each of the site-winters are defined based on the onset
date for that site-winter. The onset dates are defined as the date at which wintertime observed excess P&I mortality had been at or above a
prescribed threshold level for two continuous weeks (e.g., 0.01 deaths/100,000 people/day). Not every site-winter produced an outbreak as defined
by a particular onset threshold. Depending on the threshold level used, 1,181–1,420 epidemics were identified among 1,470 possible (30 winters each for the 48 contiguous states plus the District of Columbia). Each solid line is the averaged AH9 associated with influenza onset as defined by a different threshold mortality rate. The dashed line shows AH9 = 0. Bottom, plot of R
0(t)-anomalies using the above AH9 values. The R
0(t)-anomalies are calculated using the best combined-fit estimates of R
0max und R
0min (Table 1). The dashed line shows R0(t)9=0. R
0(t)′ = 0. doi:10.1371/journal.pbio.1000316.g002
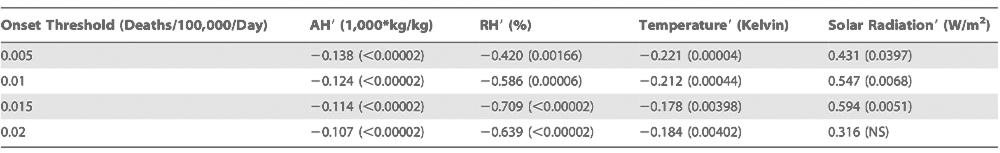
Regional differences in the association of negative AH9 with onset date are also evident. The association is strongest in the eastern US, in particular the Gulf region and the northeast (Figures S2, S3, and S4). Although the association does not reach statistical significance in much of the western US, AH9 are typically negative during the weeks prior to onset in this region as well.
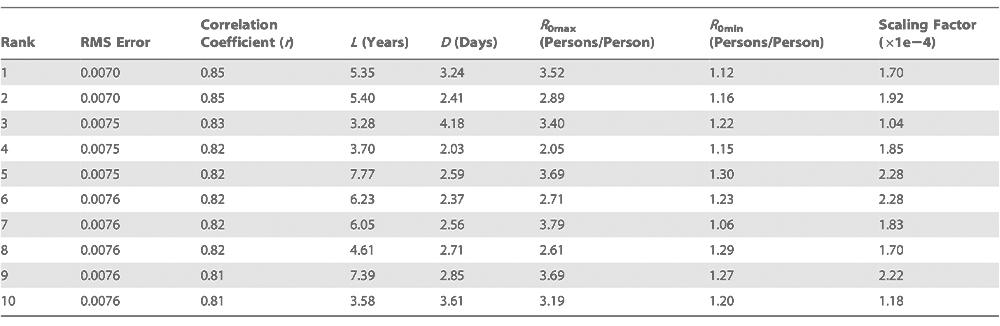
Next, we used the same approach to examine whether other potential environmental drivers of influenza are associated with wintertime influenza onset. The findings indicate that negative relative humidity (RH) and temperature anomalies, as well as positive solar insolation anomalies, are also associated with onset date (Table 1). However, the direction of the associations of the daily wintertime anomalies of solar insolation and RH with epidemic onset are contrary to the association between these environmental factors and epidemic activity at the seasonal time scale. Decreased solar insolation during the winter months is posited to increase influenza activity by decreasing host melatonin and vitamin D levels and thus host resistance [12,13]; however, our findings indicate that influenza onset is associated with increased daily solar insolation anomalies. Similarly, RH is highest in winter [11], but influenza onset is associated with low RH anomalies.
Specific weather patterns may explain the observed correlations between these meteorological anomalies and influenza onset. Anomalously low AH over the continental US is typically associated with excursions of colder air masses from the north. These air masses, which often follow a cold front, bring cloud-free skies (i.e., increased solar insolation) and reduced surface temperature and humidity levels. As the air mass moves southward, it slowly warms; however, unless it traverses a large open water source, AH does not increase substantially. As a consequence, anomalously low RH levels can develop within these air masses as well. Thus, the anomalies of solar insolation and RH could be noncausally linked with influenza outbreaks through their association with weather conditions that bring negative AH9 to a region.
Temperature and AH are strongly correlated (Table S1); both are minimal in winter when influenza transmission is maximal and have negative anomalies associated with influenza onset, tendencies which agree with the associations determined from laboratory data [11,14,15]. To establish which of these variables is most critical for onset, we rely on previous laboratory analyses exploring the impact of both environmental factors that indicate AH is the essential determinant of influenza virus survival and transmission [11]. Furthermore, AH9 is the only anomaly variable whose association with onset is significant at p < 0.00002 for all four onset threshold levels (Table 1).
Table 1
Association of daily anomalies of various environmental variables with the outbreak of influenza in the winter period 1972–2002 for the contiguous USA
In addition, it should be noted that seasonal temperature conditions are often highly managed indoors, where most of the US population spends the bulk of its time. Average daily outdoor temperatures can differ over 20 uC from winter to summer, but seasonal heating and air conditioning greatly reduce this temperature cycle indoors. In contrast, AH possesses a large seasonal cycle both outdoors and indoors [11].
Model Simulations of Influenza Seasonality
To further assess the hypothesis that AH is a fundamental driver of influenza seasonality, we examined whether a population-level model of influenza transmission forced by AH conditions could reproduce the observed seasonal patterns of P&I mortality. We simulated influenza transmission for five states representative of different climates within the US: Arizona, Florida, Illinois, New York, and Washington.
The model considers three disease classes: susceptible, infected, recovered; to integrate the impact of waning immunity following antigenic drift, we allow individuals to go back to the susceptible class at a defined rate (SIRS model). Observed 1972–2002 daily AH conditions within each state are used to modulate the basic reproductive number, R0(t), of the influenza virus, i.e., the per generation transmission rate in a fully susceptible population. These daily fluctuations of R0(t) alter the transmission probability per contact within the SIRS model and thus affect influenza transmission dynamics. The SIRS model contains four free parameters: two (R0max and R0min) that define the range of R0(t), one for the duration of immunity (D), and one for the duration of infectiousness (L).
If absolute humidity controls influenza seasonality, best-fit simulations with the AH-driven transmission model should meet the following criteria: 1) the mean annual model cycle of infection should match observations in each state; 2) these simulations should converge to similar parameter values, i.e., the virus response to AH should be consistent among states; and 3) AH modulation of transmission rates (R
0(t)) within the model must match the large range implied by the laboratory data (Figure 1).
Multiple 31-y (1972–2002) simulations were run at each of the five states with randomly chosen parameter combinations. We then compared the mean annual cycle of daily infection from each simulation with a similar average of 1972–2002 observed excess P&I mortality rates [3,4]. Best-fit model simulations at each site capture the observed seasonal cycle of influenza (Figure 3). These simulations produce not only the late-year rise in transmission and infection, but also the wintertime peak during early January, typically followed by a secondary peak during late February/early March. In both models and observations, the dual winter peaks are not typically seen in individual years; rather these epidemic trajectories reflect the averaging of individual wintertime outbreaks that peak anytime between December and April (Figure S5).
Abbildung 3
Mittlere Jahreskreisläufe der Best-Fit-SIRS-Modellsimulationen in den fünf Staaten. Hier wurden die Best-Fit-Simulationen für jeden Staat einzeln basierend auf dem RMS-Fehler nach Skalierung des 31-jährigen Mittels täglicher Infektionszahlen auf das 31-jährige Mittel beobachteter täglicher übermäßiger P&I-Sterblichkeitsraten ausgewählt. Die dicke blaue Linie zeigt die Best-Fit-Simulation; die dünneren grünen Linien zeigen die neun nächstbesten Simulationen. doi:10.1371/journal.pbio.1000316.g003
We also searched for the best-fit parameter combinations for all five sites evaluated together. The parameter combinations of these best ‘‘combined fits’’ are characterized by high R
0max (generally > 2,8), high R
0min (> 1) and low mean infectious period (2 < D < 4,2 days) (Figure S6; Table 2). Best-fit simulations at each of the five sites individually occupy a similar parameter space (Figures S7, S8, S9, S10, and S11; Table S2). In particular, these simulations converge to high R
0max, which indicates a similar response to AH variability (see Text S1).
Tabelle 2
Parameterkombinationen für die zehn Best-Fit-Simulationen an den Standorten Arizona, Florida, Illinois, New York State und Washington State
There is some correlation among SIRS model parameter values in simulations that fit the observed excess P&I mortality well. For instance, among better-fit simulations, L and D tend to be inversely related (Figures S6, S7, S8, S9, S10, and S11). In addition, broad regions of parameter space appear capable of producing highquality, low root mean square (RMS) error simulations (Figure S6). The stochastic components of the SIRS model may contribute in part to this behavior. The flat goodness-of-fit within model parameter space indicates that no one parameter combination is strictly ‘‘best,’’ rather, a range of parameter combinations may produce good simulations of influenza transmission. These parameter ranges are: L =3–8 y, D= 2–3.75 d, R
0max = 2,6–4 and R
0min = 1,05–1,30. We reran the SIRS model repeatedly sampling this approximate subset range of parameter space. Bestfit simulations from this subset range of parameter space (Table S3) were of similar quality and exhibited the same flat goodness-offit within model parameter space as the best-fit simulations presented in Table 2.
Because the SIRS model simulates only influenza-related infections, not deaths, a scaling factor is needed to compare model-simulated rates of infection with the observed excess P&I mortality rates. This scaling factor can be understood as the case fatality ratio, i.e., the probability of mortality given infection. Reassuringly, all best-fit simulations produce a scaling factor of the same order of magnitude and roughly consistent with the expected value of the case fatality ratio for P&I-related deaths (see Text S1). The model also explains regional variations in influenza dynamics. Due to the modeled nonlinear relationship between R
0(t) and AH (Figure 1C), the seasonal cycle of
0(t) is sensitive to both AH seasonal cycle amplitude and mean AH levels (Figure 1D and 1E). In Florida, mean AH levels are higher than for the other four states, but the seasonal AH cycle remains large and produces a seasonal R
0(t)-cycle of sufficient amplitude to generate an effective reproductive number R
E(t)= R
0(t) *S(t)/N, greater than 1 (Figure 1F) and organize influenza epidemics preferentially during winter.
Outbreak dynamics reinforce this phase organization in that wintertime epidemics confer immunity to a large proportion of the model population, which then reduces population-level susceptibility during the following summer when R
0(t) is low. In Arizona and Washington state, the seasonal AH cycle is less than for the other three states, but average AH levels are lower, at a range where laboratory findings indicate sensitivity to variation in AH is greater; consequently R
0(t) retains a sizeable seasonal cycle (Figure 1E). For all five states, the AH-driven seasonal variation of R
0(t) is large enough that R
(t) is strongly modulated by AH conditions and exceeds 1 during winter as outbreaks develop (Figure 1F).
The humidity-driven SIRS simulations satisfy our three criteria for supporting the hypothesis that AH controls influenza seasonality in temperate regions. The simulations produce a consistent response in the five climatologically diverse US states using similar parameter values. The large sensitivity of simulated influenza transmission to AH is consistent with the analysis of laboratory experiments that show large changes in influenza virus survival and transmission in response to AH variability (Figure 1A and 1B).
Cross-Validation of the Model Findings
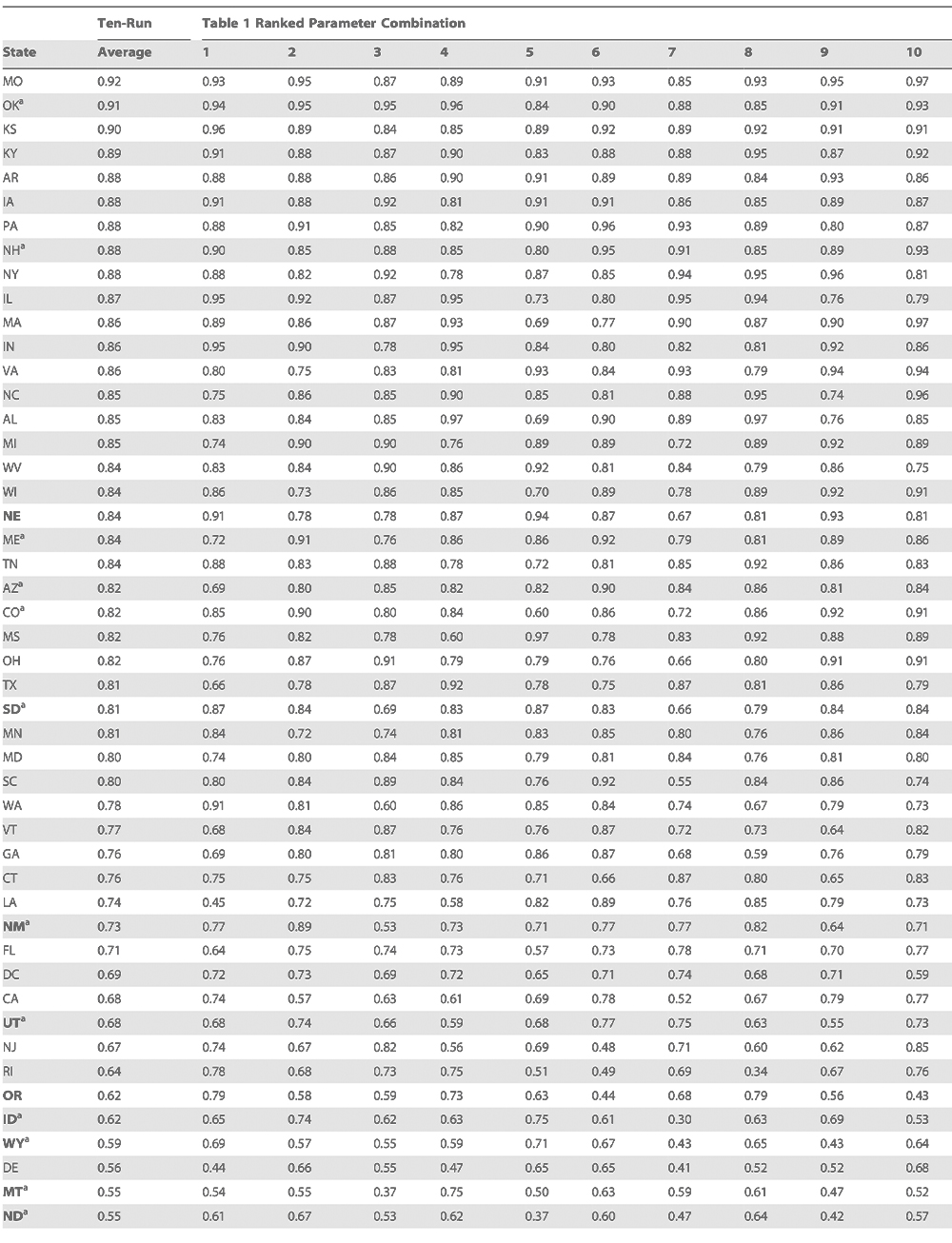
To further validate the SIRS model findings, we determined whether the best-fit simulations derived from the five selected states could reproduce the seasonal cycles of influenza elsewhere in the US. The ten best combined-fit parameter combinations (Table 2) were used to perform 31-y (1972–2002) SIRS simulations at each of the contiguous 48 states plus DC.
The results of this cross-validation demonstrate good simulations of observed excess P&I mortality for a majority of states (average r.0.7, minimum r.0.5, see Methods and Table 3).
Table 3
Correlation coefficients for the contiguous US and District of Columbia of SIRS-simulated 1972–2002 influenza incidence with 1972–2002 observed excess P&I mortality.
Some states, particularly the sparsely populated western states perform less well. These states often have low workflow [4], which may reduce the rate of introduction of the virus each winter. In addition, heterogeneous AH fields across some states (particularly large ones) create some error due to simulation with a single average statewide AH value. Thirteen states in the continental US, including Arizona, possess low workflow rates [4]. Six of these 13 states are among the ten worst cross-validation performers; such a clustering is unlikely to occur by chance alone (p <0.005). In addition, seven of the ten worst performers are states with the ten lowest population densities (p <0.0001).
Overall, the cross-validation shows that the best combined-fit parameter combinations can simulate influenza seasonality throughout the country. Future use of higher resolution AH and observed P&I data that better represent local conditions may improve these model results.
Additional SIRS Model Results
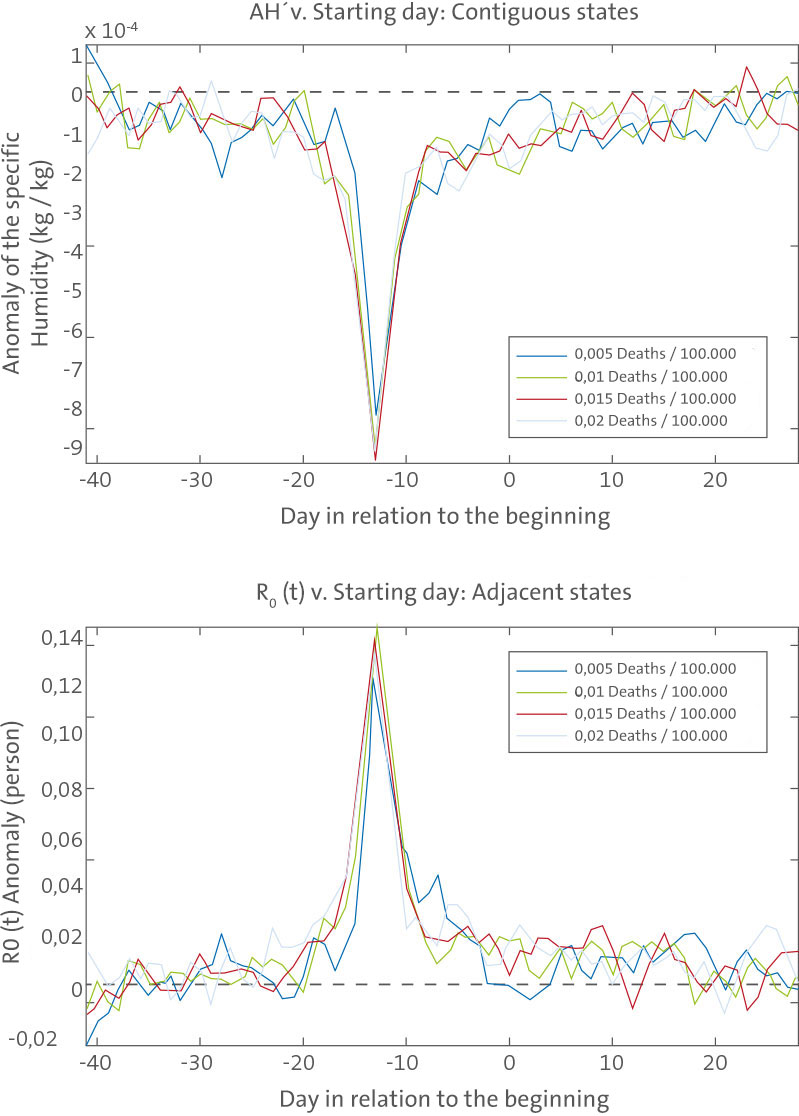
We also used SIRS model simulations to provide additional support for the association between negative AH9 and epidemic onset (Figure 2). Best-fit SIRS model runs reveal a comparable effect in which large negative AH9 develop about 2 wk prior to onset as defined by SIRS model infection rates (Figure 4). The 1- wk difference in lag between this analysis with model infection rates (2 wk) and the analysis with observed excess P&I mortality rates (3 wk) roughly corresponds to the median time from infection to mortality [16–18]. The broader peak of negative AH9 seen in Figure 2 is likely due to other, real-world factors that affect onset response and are not represented in the SIRS model (see Text S1). Finally, we examined whether the school calendar, which alters person-to-person contact rates, could provide a better simulation of seasonal influenza than AH. School holidays have been estimated to lead to changes of ,25% in influenza transmission [19] and occur during summer in the US, as well as at the end of the calendar year and again in spring.
A number of SIRS model simulations were performed that included a stepwise increase of R
0(t) during the school year (see Text S1). Simulations in which school closure was the only modulation of R
0(t) were able to generate a winter seasonal cycle of influenza; however, these simulations did not reproduce observed excess P&I mortality as well as those with AH alone (see Figures S14 and S15; Table S5; Text S1). In addition, a 40%–90% change in influenza transmission (R
0(t)) was needed to effect this seasonality (Table S5). This range of R
0(t) changes is slightly larger than the previously estimated modulation of ,25%; however, these previous estimates were derived from an age-structured population model, so direct comparison is difficult.
Discussion
Distinguishing among potential environmental drivers of influenza seasonality, such as AH, RH, temperature, solar insolation, and the school calendar, is difficult since all demonstrate a similarly strong annual periodicity. Nevertheless, our findings indicate that AH is a major (and likely the predominant) determinant of influenza seasonality due to: 1) the empirical association of negative AH9 with the onset of wintertime influenza outbreaks (Figure 2), which is statistically stronger than for RH, temperature or solar insolation (Table 1); 2) the relative consistency of the response to AH among the five states modeled in detail (i.e., similar parameter space; Table S2); and 3) the SIRS cross-validation showing that the same best-fit parameters (Table 2) can produce successful simulations of influenza seasonality throughout much of the US (Table 3).
In addition, several findings undermine the hypothesis that the association between the seasonal influenza cycle and AH is in fact due to confounding by other potential drivers. The case for solar insolation is weakened by its implausible positive association with wintertime influenza onset. Although laboratory analyses find that low RH favors influenza virus survival and transmission, RH is in fact typically incorrectly phased in the outdoor environment (i.e., maximal during winter, minimal during summer) and cannot explain peak wintertime influenza incidence.
The case for temperature is weakened by the small amplitude of its seasonal cycle in most indoor environments. Finally, reanalyses of laboratory experiments indicate that AH is the best single-variable constraint of influenza virus survival and transmission [11]; associations with temperature and RH likely merely reflect their positive covariability with AH at various time scales. Still, a role for temperature or other (possibly multiple) covariable factors cannot be entirely discounted. Further laboratory investigation is needed to determine the effects of humidity, evaporation, and temperature on virus protein structure and survival.
SIRS model simulations also indicate that although the school calendar can explain seasonal epidemic influenza, the correspondence with observations is not as good as for simulations driven by AH (Figures S14 and S15). The required increase in transmissibility during school terms is greater than estimated previously; with such large variation in transmission, inclusion of non-summer breaks creates a noticeable decline in transmission in the Christmas and spring periods that is not observed in data (see Text S1). Nonetheless, an effect of school closure on influenza transmission rates is well documented [19,20] and cannot be discounted. It is certainly possible that the effects of AH and the school calendar on influenza transmission act in concert with one another; however, our statistical and SIRS model findings indicate that AH variability provides a more parsimonious explanation for the seasonality of epidemic influenza in temperate regions, and in addition, is associated with the onset date of individual wintertime outbreaks.
The argument that AH at least partly determines influenza seasonality is supported by: 1) laboratory evidence [11]; 2) the much weaker seasonality in the tropics where humidity is high year-round, but a school calendar exists; 3) the AH9-onset analysis (Figures 2 and 4); 4) the plausibility of parameter combinations and the effect size for AH within SIRS model simulations (Figures 1 and 3; Table 2); and 5) the superior overall quality of AH-forced simulations (Figure S14) and their reduced sensitivity to stochastic processes within the SIRS model (Figure S15).
There are minor differences among the sites in the best-fit parameter values for the SIRS model (Figures S7, S8, S9, S10, and S11, and Table S2), some of which could be host mediated. For instance, Florida and New York show a tendency toward lower duration of immunity. This difference could be derived from a number of host-mediated factors specific to these states. The findings presented here do not preclude an influence of such factors on influenza transmission and seasonality. Differences in population susceptibility and infectivity (e.g., population age and general health), seasonal variations of host behavior (e.g., more time indoors in close contact during winter [19]), and host resistance (e.g., wintertime melatonin or vitamin D deficiencies [12,13]) may still affect influenza transmission rates.
Among states, there are also differences of average peak SIRSsimulated R
E(t) (Figure 1F); however, there is no systematic relationship between rates of observed excess P&I deaths and those peak R
E(t) values among these sites. For instance, Florida and New York have similar rates of observed excess mortality per 100,000 persons, but different peak R
E(t) State-to-state differences in contact rates and population age and structure, in particular the proportion of seniors, who are at highest risk of influenza-related death during seasonal epidemics, undoubtedly affect influenza infection and mortality rates, and modulate the amplitude and duration of individual outbreaks. In addition, the dominant influenza subtype is a key predictor of influenza-related mortality rate each season; A/H3N2-dominant seasons are associated with two to three times higher death rates than H1N1 and B-dominant seasons [3,4]. These other factors are not accounted for in our SIRS model; hence, there is not a good one-to-one correspondence between the average peak size of R
E(t) and rates of observed excess P&I deaths. However, within the SIRS model, a relationship between R
E(t) and simulated infection rates does exist. Among the ten best-fit simulations at each site, the average maximum
E(t) rank (from greatest to least) as New York, Illinois, Washington, Arizona, Florida. Similarly among these runs, the average maximum epidemic size ranks (from greatest to least) as New York, Washington, Illinois, Arizona, Florida. This more direct response is not unexpected; within the SIRS model, higher R
E(t) directly corresponds to greater transmission and, consequently, more rapidly developing, larger outbreaks.
It should be noted that observed excess P&I mortality is an imperfect indicator of influenza incidence, as other respiratory illnesses exhibit similar seasonal periodicities. No doubt these other diseases contribute to the seasonality of the observational time series used here (Figure 3, Figure S1). However, excess P&I mortality generally shows a strong correspondence with other indicators of influenza incidence, such as hospitalization data and laboratory notifications [4]. A clearer picture of the environmental determinants of influenza seasonality and onset will emerge when the effects of AH and other environmental variables on these potentially confounding, seasonal respiratory pathogens are also elucidated.
The initial evidence demonstrating that AH affects influenza virus survival and transmission was derived from laboratory experiments studying the airborne transmission of influenza; however, our SIRS model uses no specific mode of transmission. Thus, other modes of transmission, in particular indirect transmission via fomites, if similarly affected by AH, might also have a role determining the seasonality of influenza in temperate regions. In addition, the SIRS model is highly idealized and fails to represent many factors in the real world that can affect transmission rates, including clustered populations, structured interactions, variation in host infectiousness, and multiple influenza strains conferring various levels of cross-immunity.
Figure 4
AH9 associated with SIRS simulated influenza onset. Top, plots of average AH9 associated with wintertime influenza onset for the ten best-fit SIRS model simulations at the five state sites (Arizona, Florida, Illinois, New York, and Washington). The onset dates are defined as the date on which wintertime infection rates have been at or above a prescribed level for two continuous weeks (e.g., 50 infections/100,000 people/day). Each solid line is the averaged AH9 associated with influenza onset as defined by a different threshold infection rate. The dashed line shows AH9=0. Bottom, plots of R
0(t) anomalies using the AH9 values. The R
0(t) anomalies are calculated using the parameters R
0max and R
0min from each best-fit simulation (Table S3). The dashed line shows R
0(t)′ = 0.
doi:10.1371/journal.pbio.1000316.g004
Future work could incorporate these effects into a more realistic influenza model that also accounts for the effects of AH. Such efforts would also enable better discrimination between the effects of school terms and AH. Also, the effects of AH on influenza transmission should be incorporated into models accounting for travel and workflow [4,21,22] to explain the seasonal geographic spread of influenza.
The analyses presented here need to be extended elsewhere, including the tropics, where AH is high year-round and the seasonality of influenza is often less clearly defined. High AH does not preclude but merely reduces influenza virus survival and transmission, so it is possible a role for AH also exists in the tropics.
However, the findings presented here suggest that R
0(t) would be less sensitive to AH variability in areas of very high year-round AH, such as the tropics, which may allow for other, possibly hostmediated, factors to play a more predominant role in generating seasonal variability in influenza incidence. Laboratory studies provided the initial evidence that AH may determine the seasonality of influenza in temperate regions [11]. The model and statistical results presented here indicate that the effects of AH observed in the laboratory are sufficient to explain patterns observed at the population level and illustrate the power of epidemiological modeling rooted in individual-level experiments. The results indicate that AH affects both the seasonality of influenza incidence and the timing of individual wintertime influenza outbreaks in temperate regions. The association of negative AH9 with wintertime influenza outbreak onset is remarkable given the noise in the data and suggests that skillful, short-term probabilistic forecasts of epidemic influenza could be developed.
Methods
SIRS Model and Methods
The SIRS model equations are:
where S is the number of susceptible people in the population, t is time in years, N is the population size, I is the number of infectious people, N2S2I is the number of resistant individuals, b(t) is the contact rate at time t, L is the average duration of immunity in years, and D is the mean infectious period in years. The basic reproductive number at time t is R
0(t)= ß(t)D.
Observed AH conditions were derived from National Centers for Environmental Prediction–National Center for Atmospheric Research (NCEP-NCAR) reanalysis [23]. For each state (Arizona, Florida, Illinois, New York, and Washington), a daily 1972–2002 time series of 2-m above-ground specific humidity, q(t), was constructed by averaging all grid cells with $10% of their area within that state. The equation relating q(t) to R0(t) uses an exponential functional form similar to the relationships between AH and both influenza virus survival and transmission, derived from laboratory experiments (Figure 1):
where a = - 180, b = log(R
0max - R
0min), R
0max is the maximum daily basic reproductive number, and R
0min is the minimum daily basic reproductive number. The value of a is estimated from the laboratory regression of influenza virus survival upon AH (Figure 1A). Equation 4 dictates that R
0(t)= R
0max, when q(t) = 0 kg/kg and that R
(t) approaches R
0min asymptotically as q(t) increases. For simulations, we use a stochastic Markov chain formulation in which individuals are treated as discrete entities, and transitions between model states (i.e., susceptible, infected, recovered) are determined by random draws corresponding to rates determined from Equations 2 and 3. Using the daily time series of q(t), which alters R
0(t) daily influenza transmission was simulated for each of the five state sites during 1972–2002 for a model population of 500,000 individuals. Initial conditions included 50,000 susceptible persons and 100 infected persons; however, results were not sensitive to these numbers. Simulations were performed with daily random seeding of infected individuals (i.e., each day there is a 10% probability that a single susceptible individual becomes infected), meant to represent reintroduction of the virus in the model domain due to travel. The model is perfectly mixed and simulations were performed with two influenza virus subtypes: AH1N1/ B and A-H3N2 (see Figures S12 and S13). No cross immunity was conferred within the model between these virus subtypes.
Each year, beginning in May, the random seeding of infectious individuals in the population (representing emigration/ travel) is fixed to the dominant recorded subtype for the US (i.e., either A-H1N1/B or A-H3N2), based on Centers for Disease Control/Morbidity and Mortality Weekly Report (CDC/ MMWR) laboratory and antigenic surveillance data [3,4]. Five thousand simulations at each site were run using combinations of the four model parameters: R
0max, R
0min, D and L chosen using a Latin hypercube sampling structure with uniform distribution. The ranges of these parameters were specified to reflect known influenza dynamics (see Table S4, Text S1). Estimates of R
0, derived or used by many authors range from 1.3 to 3 [16,21,22,24–27]. To effect this range, given Equation 3 and variations of q(t), we used an R
0max ranging from 1.3 to 4.
R
0min provides the R
0 below which the model cannot fall. This minimum recognizes that other modes of influenza transmission exist that may not be modulated by absolute humidity. R
0min values range from 0.8 to 1.3. Per Equation 4, decreasing humidity increases R
0(t) The range of this nonlinear increase is set by the randomly chosen R
0max and R
0min parameters. Estimates of D range from 2 to 7 d [16,25], and estimates of L range from 2 to 10 y. Both influenza virus subtypes used the same four randomly chosen parameters during each simulation, though results were similar when each subtype was assigned different parameters (eight in total). The quality of each simulation at each site was evaluated based on RMS error with observed excess P&I mortality [3,4] (Figure S1), lagged 2 wk. The lag accounts for mean time from infection to mortality. Prior to determining the RMS error, each model run was scaled to enable comparison of simulated infections with observed mortality rates (see Text S1).
Prior to the model cross-validation throughout the contiguous US, we first determined the effect that stochasticity within each SIRS simulation has on the quality of fit with observations. We reran the ten best common-fit parameter combinations 100 times each for New York, each time with a different random seeding, and found that correlations to observed P&I mortality ranged from r =0.65 to r= 0.97 with an average of r= 0.87. In contrast, multiple simulations with the 4,500th best parameter combination, out of 5,000, produced correlations that ranged from r =20.18 to r =0.34. Thus, random seeding within a particular model run produces a range of correlation coefficient outcomes; however, good model runs should produce high positive correlation with observations (average r>0.7, minimum r>0.5).